Макромолекулярная структура ДНК
В 1953 г. Уотсон и Крик, опираясь на известные данные о конформаци нуклеозидных остатков, о характере межнуклеотидной связи в ДНК и закономерности нуклеотидного состава ДНК (правила Чаргаффа), расшифровали рентгенограммы паракристаллической формы ДНК [так называемой В-формы, образующейся при влажности выше 80% и при высокой концентрации противоионов (Li+) в образце]. Согласно их модели, молекула ДНК представляет собой правильную спираль, образованную двумя полидезоксирибонуклеотидными цепями, закрученными относительно друг друга и вокруг общей оси. Диаметр спирали практически постоянен вдоль всей ее длины и равен 1,8 нм (18 А).
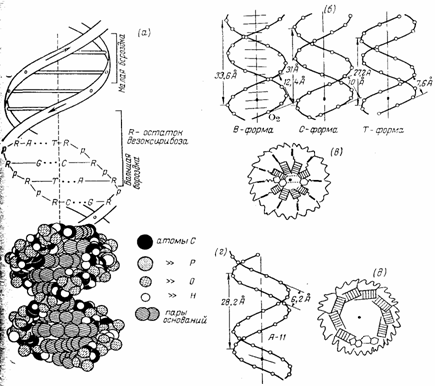
Макромолекулярная структура ДНК.
(а)—Модель Уотсона — Крика;
(6)—параметры спиралей В-, С- и Т-форм ДНК (проекции перпендикулярно оси спирали);
(в)—поперечный разрез спирали ДНК в В-форме (заштрихованные прямоугольники изображают пары оснований);
(г)—параметры спирали ДНК в А-форме;
(д)—поперечный разрез спирали ДНК в А-форме.
Длина витка спирали, который соответствует ее периоду идентичности, составляет 3,37 нм (33,7 А). На один виток спирали приходится 10 остатков оснований в одной цепи. Расстояние между плоскостями оснований равно, таким образом, примерно 0,34 нм (3,4 А). Плоскости остатков оснований перпендикулярны длинной оси спирали. Плоскости углеводных остатков несколько отклоняются от этой оси (первоначально Уотсон и .Крик предположили, что они параллельны ей).
Из рисунка видно, что углеводофосфатный остов молекулы обращен наружу. Спираль закручена таким образом, что на ее поверхности можно выделить две различные по размерам бороздки (их часто называют также желобками) — большую, шириной примерно 2,2 нм (22 А), и малую —шириной около 1,2 нм (12А). Спираль — правовращающая. Полидезоксирибонуклеотидные цепи в ней антипараллельны: это означает, что если мы будем двигаться вдоль длинной оси спирали от одного ее конца к другому, то в одной цепи мы будем проходить фосфодиэфирные связи в направлении 3'à5', а в другой — в направлении 5'à3'. Иными словами, на каждом из концов линейной молекулы ДНК расположены 5'-конец одной и 3'-конец другой цепи.
Регулярность спирали требует, чтобы против остатка пуринового основания в одной цепи находился остаток пиримидинового основания в другой цепи. Как уже подчеркивалось, это требование реализуется в виде принципа образования комплементарных пар оснований, т. е. остаткам аденина и гуанина в одной цепи соответствуют остатки тимина и цитозина в другой цепи (и наоборот).
Таким образом, последовательность нуклеотидов в одной цепи молекулы ДНК предопределяет нуклеотидную последовательность другой цепи.
Этот принцип является главным следствием модели Уотсона и Крика, поскольку он в удивительно простых химических терминах объясняет основное функциональное назначение ДНК — быть хранителем генетической информации.
Заканчивая рассмотрение модели Уотсона и Крика, остается добавить, что соседние пары остатков оснований в ДНК, находящейся в В-форме, повернуты друг относительно друга на 36° (угол между прямыми, соединяющими атомы С1' в соседних комплементарных парах).
Смотрите также
Барий. Свойства, получение, распространение
Тяжелый шпат, BaSO4 ,
был первым известным соединением барин. Его открыл в начале XVII в. итальянский
алхимик Касциароло. Он же установил, что этот минерал после сильного нагревания
с углем ...
Кремний, полученный с использованием геттерирования расплава
В бездефектной технологии
изготовления ИС для уменьшения влияния термодефектов используются методы
пассивного геттерирования примесей в пластинах. К таким методам относятся
"внешнее г ...
Четвертая группа периодической системы.
По электронным структурам нейтральных атомов к углероду и
кремнию примыкают германий и его аналоги. Максимальная валентность
этих элементов, как по отдаче, так и по присоединению электро ...